
Data availability
The atomic coordinates included in this study have been deposited in the PDB with the following accession codes: 8KAG ref. 50, 8KAH, 8KAI, 8KAJ, 8KAK, 8KAL and 8KAM. PDB IDs 4OO8 and 5U33 were used for Fig. 1a and Extended Data Fig. 9a. PDB ID 7Z4J was used for Extended Data Fig. 8a,d. PDB ID 4ZT0 was used for Extended Data Fig. 9a,b. Source data are provided with this paper.
References
Makarova, K. S., Zhang, F. & Koonin, E. V. SnapShot: class 2 CRISPR–Cas systems. Cell 168, 328–328.e1 (2017).
Article
CAS
PubMed
Google Scholar
Cong, L. et al. Multiplex genome engineering using CRISPR/Cas systems. Science 339, 819–823 (2013).
Article
CAS
PubMed
PubMed Central
Google Scholar
Li, J. F. et al. Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nat. Biotechnol. 31, 688–691 (2013).
Article
CAS
PubMed
PubMed Central
Google Scholar
Nekrasov, V., Staskawicz, B., Weigel, D., Jones, J. D. & Kamoun, S. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 31, 691–693 (2013).
Article
CAS
PubMed
Google Scholar
Jiang, W., Bikard, D., Cox, D., Zhang, F. & Marraffini, L. A. RNA-guided editing of bacterial genomes using CRISPR–Cas systems. Nat. Biotechnol. 31, 233–239 (2013).
Article
CAS
PubMed
PubMed Central
Google Scholar
Zetsche, B. et al. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR–Cas system. Cell 163, 759–771 (2015).
Article
CAS
PubMed
PubMed Central
Google Scholar
Fozouni, P. et al. Amplification-free detection of SARS-CoV-2 with CRISPR–Cas13a and mobile phone microscopy. Cell 184, 323–333 (2021).
Article
CAS
PubMed
Google Scholar
Broughton, J. P. et al. CRISPR–Cas12-based detection of SARS-CoV-2. Nat. Biotechnol. 38, 870–874 (2020).
Article
CAS
PubMed
PubMed Central
Google Scholar
Shmakov, S. et al. Discovery and functional characterization of diverse class 2 CRISPR–Cas systems. Mol. Cell 60, 385–397 (2015).
Article
CAS
PubMed
PubMed Central
Google Scholar
Yamano, T. et al. Crystal structure of Cpf1 in complex with guide RNA and target DNA. Cell 165, 949–962 (2016).
Article
CAS
PubMed
PubMed Central
Google Scholar
Gao, P., Yang, H., Rajashankar, K. R., Huang, Z. & Patel, D. J. Type V CRISPR–Cas Cpf1 endonuclease employs a unique mechanism for crRNA-mediated target DNA recognition. Cell Res. 26, 901–913 (2016).
Article
CAS
PubMed
PubMed Central
Google Scholar
Dong, D. et al. The crystal structure of Cpf1 in complex with CRISPR RNA. Nature 532, 522–526 (2016).
Article
CAS
PubMed
Google Scholar
Liu, L. et al. Two distant catalytic sites are responsible for C2c2 RNase activities. Cell 168, 121–134 (2017).
Article
CAS
PubMed
Google Scholar
Swarts, D. C. & Jinek, M. Mechanistic insights into the cis– and trans-acting DNase activities of Cas12a. Mol. Cell 73, 589–600 (2019).
Article
CAS
PubMed
PubMed Central
Google Scholar
Abudayyeh, O. O. et al. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science 353, aaf5573 (2016).
Article
PubMed
PubMed Central
Google Scholar
Li, S. Y. et al. CRISPR–Cas12a has both cis– and trans-cleavage activities on single-stranded DNA. Cell Res. 28, 491–493 (2018).
Article
CAS
PubMed
PubMed Central
Google Scholar
Chen, J. S. et al. CRISPR–Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 360, 436–439 (2018).
Article
CAS
PubMed
PubMed Central
Google Scholar
East-Seletsky, A. et al. Two distinct RNase activities of CRISPR–C2c2 enable guide-RNA processing and RNA detection. Nature 538, 270–273 (2016).
Article
CAS
PubMed
PubMed Central
Google Scholar
Gootenberg, J. S. et al. Nucleic acid detection with CRISPR–Cas13a/C2c2. Science 356, 438–442 (2017).
Article
CAS
PubMed
PubMed Central
Google Scholar
Gootenberg, J. S. et al. Multiplexed and portable nucleic acid detection platform with Cas13, Cas12a, and Csm6. Science 360, 439–444 (2018).
Article
CAS
PubMed
PubMed Central
Google Scholar
Myhrvold, C. et al. Field-deployable viral diagnostics using CRISPR–Cas13. Science 360, 444–448 (2018).
Article
CAS
PubMed
PubMed Central
Google Scholar
Rananaware, S. R. et al. Programmable RNA detection with CRISPR–Cas12a. Nat. Commun. 14, 5409 (2023).
Article
CAS
PubMed
PubMed Central
Google Scholar
Teng, F. et al. CDetection: CRISPR–Cas12b-based DNA detection with sub-attomolar sensitivity and single-base specificity. Genome Biol. 20, 132 (2019).
Article
PubMed
PubMed Central
Google Scholar
Sapranauskas, R. et al. The Streptococcus thermophilus CRISPR/Cas system provides immunity in Escherichia coli. Nucleic Acids Res. 39, 9275–9282 (2011).
Article
CAS
PubMed
PubMed Central
Google Scholar
Jinek, M. et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337, 816–821 (2012).
Article
CAS
PubMed
PubMed Central
Google Scholar
Gasiunas, G., Barrangou, R., Horvath, P. & Siksnys, V. Cas9–crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc. Natl Acad. Sci. USA 109, E2579–E2586 (2012).
Article
CAS
PubMed
PubMed Central
Google Scholar
Nishimasu, H. et al. Crystal structure of Cas9 in complex with guide RNA and target DNA. Cell 156, 935–949 (2014).
Article
CAS
PubMed
PubMed Central
Google Scholar
Yang, H., Gao, P., Rajashankar, K. R. & Patel, D. J. PAM-dependent target DNA recognition and cleavage by C2c1 CRISPR–Cas endonuclease. Cell 167, 1814–1828 (2016).
Article
CAS
PubMed
PubMed Central
Google Scholar
Pacesa, M. et al. R-loop formation and conformational activation mechanisms of Cas9. Nature 609, 191–196 (2022).
Article
CAS
PubMed
PubMed Central
Google Scholar
Zhu, X. et al. Cryo-EM structures reveal coordinated domain motions that govern DNA cleavage by Cas9. Nat. Struct. Mol. Biol. 26, 679–685 (2019).
Article
CAS
PubMed
PubMed Central
Google Scholar
Nierzwicki, L. et al. Principles of target DNA cleavage and the role of Mg2+ in the catalysis of CRISPR–Cas9. Nat. Catal. 5, 912–922 (2022).
Article
CAS
PubMed
PubMed Central
Google Scholar
Jiang, F., Zhou, K., Ma, L., Gressel, S. & Doudna, J. A. A Cas9-guide RNA complex preorganized for target DNA recognition. Science 348, 1477–1481 (2015).
Article
CAS
PubMed
Google Scholar
Reita, D. et al. Molecular mechanism of EGFR-TKI resistance in EGFR-mutated non-small cell lung cancer: application to biological diagnostic and monitoring. Cancers 13, 4926 (2021).
Article
CAS
PubMed
PubMed Central
Google Scholar
Harrington, L. B. et al. Programmed DNA destruction by miniature CRISPR–Cas14 enzymes. Science 362, 839–842 (2018).
Article
CAS
PubMed
PubMed Central
Google Scholar
Ackerman, C. M. et al. Massively multiplexed nucleic acid detection with Cas13. Nature 582, 277–282 (2020).
Article
CAS
PubMed
PubMed Central
Google Scholar
Bravo, J. P. K. et al. Structural basis for mismatch surveillance by CRISPR–Cas9. Nature 603, 343–347 (2022).
Article
CAS
PubMed
PubMed Central
Google Scholar
Stephenson, A. A., Raper, A. T. & Suo, Z. C. Bidirectional degradation of DNA cleavage products catalyzed by CRISPR/Cas9. J. Am. Chem. Soc. 140, 3743–3750 (2018).
Article
CAS
PubMed
Google Scholar
Huyke, D. A. et al. Enzyme kinetics and detector sensitivity determine limits of detection of amplification-free CRISPR–Cas12 and CRISPR–Cas13 diagnostics. Anal. Chem. 94, 9826–9834 (2022).
Article
CAS
PubMed
Google Scholar
Dmytrenko, O. et al. Cas12a2 elicits abortive infection through RNA-triggered destruction of dsDNA. Nature 613, 588–594 (2023).
Article
CAS
PubMed
PubMed Central
Google Scholar
Jiao, C. et al. Noncanonical crRNAs derived from host transcripts enable multiplexable RNA detection by Cas9. Science 372, 941–948 (2021).
Article
CAS
PubMed
PubMed Central
Google Scholar
Liu, Y. et al. Reprogrammed tracrRNAs enable repurposing of RNAs as crRNAs and sequence-specific RNA biosensors. Nat. Commun. 13, 1937 (2022).
Article
CAS
PubMed
PubMed Central
Google Scholar
Marquez-Costa, R. et al. Multiplexable and biocomputational virus detection by CRISPR–Cas9-mediated strand displacement. Anal. Chem. 95, 9564–9574 (2023).
Article
CAS
PubMed
PubMed Central
Google Scholar
Pardee, K. et al. Rapid, low-cost detection of Zika virus using programmable biomolecular components. Cell 165, 1255–1266 (2016).
Article
CAS
PubMed
Google Scholar
Liu, S., Rao, X., Zhao, R. & Han, W. The trans DNA cleavage activity of Cas12a provides no detectable immunity against plasmid or phage. Front. Genome Ed. 4, 929929 (2022).
Article
PubMed
PubMed Central
Google Scholar
Marino, N. D., Pinilla-Redondo, R. & Bondy-Denomy, J. CRISPR–Cas12a targeting of ssDNA plays no detectable role in immunity. Nucleic Acids Res. 50, 6414–6422 (2022).
Article
CAS
PubMed
PubMed Central
Google Scholar
Otwinowski, Z. & Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 276, 307–326 (1997).
Article
CAS
PubMed
Google Scholar
Mccoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007).
Article
CAS
PubMed
PubMed Central
Google Scholar
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 66, 486–501 (2010).
Article
CAS
PubMed
PubMed Central
Google Scholar
Adams, P. D. et al. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr. D Biol. Crystallogr. 58, 1948–1954 (2002).
Article
PubMed
Google Scholar
Chen, Y., Chen, J. & Liu, L. Crystal structure of SpyCas9 in complex with sgRNA and target RNA. Protein Data Bank https://doi.org/10.2210/pdb8kag/pdb (2023).
Download references
Acknowledgements
We are grateful to the staff of the BL-18U1, BL-19U1, BL-10U2 and BL-02U1 beamlines at the National Center for Protein Sciences Shanghai at Shanghai Synchrotron Radiation Facility. We thank W. Han for providing the plasmids and C. Wu, Y. Wu, J. Hu and W. Zheng for technical support. This work was supported by grants from the National Natural Science Foundation of China (32022047), the Natural Science Foundation of Fujian Province of China (2023J01023 and 2023J05008) and the Science and Technology Project of Xiamen City (3502Z20227020).
Author information
Author notes
These authors contributed equally: Jiyun Chen, Ying Chen, Linglong Huang.
Authors and Affiliations
State Key Laboratory of Cellular Stress Biology, School of Life Sciences, Faculty of Medicine and Life Sciences, Xiamen University, Xiamen, China
Jiyun Chen, Ying Chen, Linglong Huang, Xiaofeng Lin, Hong Chen, Wenwen Xiang & Liang Liu
Contributions
J.C., Y.C. and L.H. expressed and purified the proteins and grew crystals. H.C., X.L., Y.C., W.X. and L.L. collected X-ray diffraction data. L.L. solved the structures. J.C., Y.C., L.H., X.L. and H.C. performed all of the cloning and the biochemical assays. J.C. and L.L. prepared the figures. L.L. wrote the manuscript and supervised all of the research.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Biotechnology thanks Piyush Jain and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
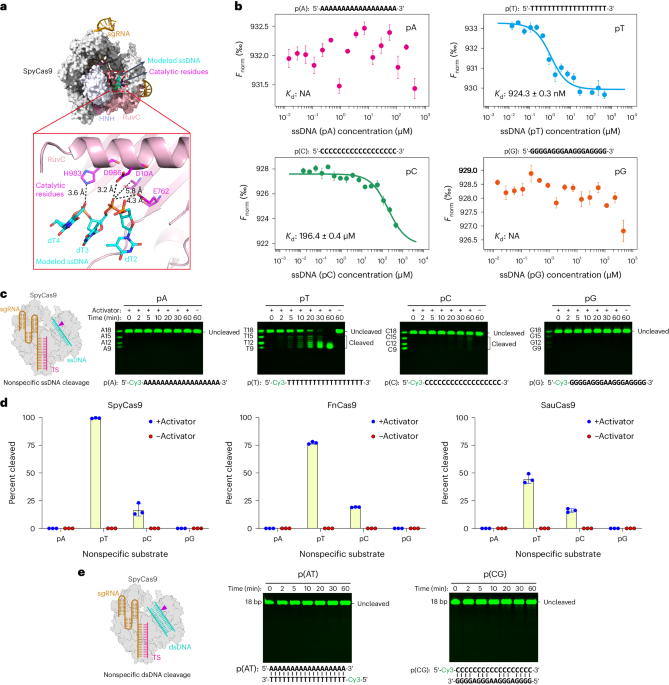
Extended Data Fig. 1 Cas9 shows similar trans-cleavage activity to Cas12a.
a-c, Denaturing gel showing the trans-cleavage of Cy3-labeled homopolymeric ssDNAs (poly A, poly T, poly C and poly G) by FnCas12a (a), FnCas9 (b), and SauCas9 (c) in the presence of a target ssDNA activator. All assays in Extended Data Fig. 1 were repeated three times independently with similar results.
Source data
Extended Data Fig. 2 Kinetic analysis of crRNA-tracrRNA and sgRNA guided trans-cleavage.
a, Representative ssDNA-FQ standard curve for kinetic parameters determination. b-e, Plots of fluorescence intensity versus time for crRNA-tracrRNA (b) or sgRNA (d) guided trans-cleavage, using target ssDNA01 activator and increasing ssDNA-FQ substrate concentrations. Michaelis-Menten fits for crRNA-tracrRNA (c) or sgRNA (e) guided ssDNA-FQ trans-cleavage using target ssDNA01 activator. The concentrations of SpyCas9 used in assays were listed in graph. f-g, Comparison of the kinetic parameters of crRNA-tracrRNA and sgRNA guided trans-cleavage with ss/dsDNA02 (f) or ss/dsDNA03 (g) activator. kcat/Km values are shown as mean ± s.d. (n = 3 independent experiments). Statistical analysis was by two-tailed t test.
Source data
Extended Data Fig. 3 Trans-cleavage of non-specific ssDNA directed by crRNA and tracrRNA.
a, Comparison of sgRNA and crRNA-tracrRNA guided FnCas9 trans-cleavage on M13 phage ssDNA substrate using a target ssDNA activator. There is no sequence homology between the M13 ssDNA and the target ssDNA. b, crRNA-tracrRNA guided SpyCas9 trans-cleavage assays using 5′-Cy5 labeled non-specific dsDNA-1 substrate with random sequence. c, crRNA-tracrRNA guided SpyCas9 trans-cleavage assays using 5′-Cy5 labeled non-specific ssDNA-2 (left) and 3′-Cy5 labeled non-specific ssDNA-3 (right) substrates with random sequences. Assays in Extended Data Fig. 3a–c were repeated three times independently with similar results. d, Schematic representation of an assay to explore the effect of the trans-cleavage activity of Cas9 on bacterial immunity. e, SpyCas9 can target and delete the pTarget plasmid in Escherichia coli. The pCas9 and pTarget plasmids contain the Chl-resistant gene and Amp-resistant gene, respectively. The cells were serially diluted and dropped onto plates containing indicated antibiotics. Control: The guide sequence of crRNA is not complementary to the target sequence (pTarget). Targeting: The guide sequence of crRNA is complementary to the target sequence (pTarget). f, The trans-cleavage activity of SpyCas9 does not prevent plague formation. Ten-fold serial dilutions of M13 phage were dropped onto bacterial lawns. g, The titers of M13 phage after infecting Escherichia coli. PFU values are shown as mean ± s.d. (n = 3 independent experiments). Statistical analysis was by two-tailed t test.
Source data
Extended Data Fig. 4 Effects of substrate DNA sequence and secondary structure on trans-cleavage activity.
a, Schematic of long non-specific ssDNA substrates. The DNA sequences of LT2, LT3 and LT4 are derived from the genomes of Microviridae sp. isolate SD_SC_96 (Genbank: MH572378.1), Microviridae sp. isolate ctzV77 (Genbank: BK042979.1), and Microvirus mar36 (Genbank: MZ089782.1), respectively. The single-stranded poly T regions of substrates are colored blue. Poly T to poly A mutations are induced in LA2, LA3 and LA4 corresponding to LT2, LT3 and LT4, respectively. Poly A regions of substrates are colored green. Poly T to poly TC mutation is induced in LTC4 corresponding to LT4. Poly T to poly TA mutation is induced in LTA4 corresponding to LT4. b-c, Testing the crRNA-tracrRNA induced trans-cleavage activity of SpyCas9 (b) or FnCas9 (c) on long non-specific ssDNA substrates containing poly T, poly A, poly TC or poly TA sequences. All data in Extended Data Fig. 4b,c are shown as mean ± s.d. (n = 3 independent experiments).
Source data
Extended Data Fig. 5 The RuvC domain is responsible for the trans-cleavage activity of SpyCas9.
a-c, Denaturing gel showing target dsDNA cis-cleavage (a), target ssDNA cis-cleavage (b), and non-specific ssDNA trans-cleavage (c) by SpyCas9 wild-type (WT), RuvC-dead mutant (D10A) or HNH-dead mutant (H840A). dsDNA and ssDNA substrates were labeled with Cy5 or Cy3. Assays in Extended Data Fig. 5 were repeated three times independently with similar results.
Source data
Extended Data Fig. 6 Substrate sequence preference of Cas9 cis-cleavage.
a, Denaturing gel showing the cleavage of target strand (TS) of target dsDNA. The target strands of target dsDNAs were all labeled with 6-FAM. Poly T, poly A, poly G and poly C in the target strands are highlighted in green, magenta, red and limon, respectively. Cleavage site of target strand is indicated by blue triangle. b, Denaturing gel showing the cleavage of non-target strand (NTS) of target dsDNA. The non-target strands of target dsDNAs were all labeled with 6-FAM. Poly T, poly A, poly G and poly C in the non-target strands are highlighted in green, magenta, red and limon, respectively. Cleavage site of non-target strand is indicated by blue triangle. c, (Top) Sequences of target dsDNA plasmids and complementary guide RNAs used in reactions. Poly T and poly G in the non-target strands are highlighted in green and magenta, respectively. Cleavage site of non-target strand is indicated by blue triangle. (Bottom) Agarose gel analysis showing the DNA cleavage by SpyCas9 with indicated target dsDNA. Assays in Extended Data Fig. 6 were repeated three times independently with similar results.
Source data
Extended Data Fig. 7 The formation of an 18-nt guide-target DNA duplex is essential for the complete activation of Cas9.
a-c, (Top) Fluorescence time-course assays showing the sensitivity of SpyCas9-catalyzed trans-cleavage using 13-nt (a), 11-nt (b), and 8-nt (c) ssDNA-FQ substrates and increasing target activator concentrations. (Bottom) Comparison of slopes of curves within 10-25 min from the top time-course graphs. The slope was calculated by simple linear regression. Data are shown as mean ± s.d. (n = 3 independent experiments). Statistical analysis was by two-tailed t test. d, (Top) Comparison of trans-cleavage rates within 10-20 min for SpyCas9 using dsDNA activators with indicated mismatches. (Bottom) Sequences of dsDNA activators and the complementary guide RNAs used in reactions. Data are shown as mean ± s.d. (n = 3 independent experiments). e-f, Time-course assays (e) and maximum fluorescence signals (f) showing that the DNA target length affects the trans-cleavage activity of SpyCas9. Data are shown as mean ± s.d. (n = 3 independent experiments).
Source data
Extended Data Fig. 8 Structural differences between SpyCas9-sgRNA-NTS_TS and SpyCas9-crRNA-tracrRNA-NTS_TS complexes.
a, Structures of SpyCas9-sgRNA-NTS_TS complexes with the 16-nt (top), 17-nt (middle first) and 18-nt (middle second) target strands in same orientation. (Bottom) The structure of SpyCas9-sgRNA-NTS_TS18 complex in catalytic state (PDB: 7Z4J). b, Structures of SpyCas9-crRNA-tracrRNA-NTS_TS complexes with the 16-nt (upper), 17-nt (middle) and 18-nt (bottom) target strands. c, SEC purification of SpyCas9-sgRNA-NTS_TS18 and SpyCas9-crRNA-tracrRNA-NTS_TS18 complexes. SEC assays were repeated three times independently with similar results. d, Structural comparison of the REC1 and HNH domains, as well as the sgRNA and target DNA in the checkpoint and catalytic state complexes. The checkpoint complex structure is shown in gray. In addition to the magenta core of the REC1 domain, the catalytic complex structure is colored according to a.
Source data
Extended Data Fig. 9 Comparison of structures of SpyCas9-sgRNA-target ssDNA and SpyCas9-sgRNA-target ssRNA.
a, Structural comparison between the SpyCas9-sgRNA-target ssDNA (PDB: 4OO8) complex and the SpyCas9-sgRNA (PDB: 4ZT0) complex. Vector length correlates with the domain motion scale. b, Structural comparison between the SpyCas9-sgRNA-target ssRNA complex and the SpyCas9-sgRNA complex (PDB: 4ZT0). c, Superposition of the SpyCas9-sgRNA-target ssRNA complex onto the SpyCas9-sgRNA-target ssDNA complex. The structure of ssDNA bound state is colored in silver. Protein domains of SpyCas9-sgRNA-target ssRNA are colored according to Fig. 4g. d, Structural overlay of the little variation domains in ssRNA bound state and ssDNA bound state complexes. e, Structural overlay of the domains that underwent marked conformational change in ssRNA bound state and ssDNA bound state complexes. f, Structural overlay of nucleic acids in ssRNA bound state and ssDNA bound state complexes. g, Superposition of the guide RNA (orange)-target ssRNA (megenta) duplex onto an A-form RNA duplex (gray).
Extended Data Fig. 10 Applications of Cas9-detection.
a-b, Fluorescence time-course assays showing the sensitivity of RACD (a) and DACD (b) to mpox (CB) strain. c, The net fluorescence intensity showing the specificity of DACD to mpox (CB) strain. Data are shown as mean ± s.d. (n = 3 independent experiments). Statistical analysis was by two-tailed t test. d, Comparison of trans-cleavage activity of SpyCas9 using dsDNA activators with indicated mismatches (Top). Sequences of the dsDNA activators and the complementary guide RNAs used in reactions (bottom). Data are shown as mean ± s.d. (n = 3 independent experiments). e-f, Fluorescence time-course assays showing the sensitivity of RACD (e) and DACD (f) to EGFR T790M mutant. g, Sensitivity analysis of DACD for detecting the EGFR T790M mutant minor allele in mock cell-free DNA samples. Data are shown as mean ± s.d. (n = 3 independent experiments). Statistical analysis was by two-tailed t test. h, Rader charts compare the features related to the application of trans-cleavage activity of Cas9 (left), Cas12a (middle) and Cas13a (right) in nucleic acid detection. Each of the 7 features form individual axes varying from 0 (without the feature) to 100 (with the feature). The vertexes of the smaller gray heptagon indicate 50 (feature can be realized by artificial modification). The value of each feature is depicted by a small circle, except that the 0 point is not displayed. The area bounded by connecting the data values for each axis is filled with the corresponding color (Cas9: purple, Cas12a: green, Cas13a: blue).
Source data
Supplementary information
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Reprints and permissions
About this article


Cite this article
Chen, J., Chen, Y., Huang, L. et al. Trans-nuclease activity of Cas9 activated by DNA or RNA target binding.
Nat Biotechnol (2024). https://doi.org/10.1038/s41587-024-02255-7
Download citation
Received: 18 September 2023
Accepted: 18 April 2024
Published: 29 May 2024
DOI: https://doi.org/10.1038/s41587-024-02255-7
>>> Read full article>>>
Copyright for syndicated content belongs to the linked Source : Nature.com – https://www.nature.com/articles/s41587-024-02255-7